biostar
Animateur Des Topics


 Nombre de messages : 982 Nombre de messages : 982
Age : 34
Localisation : kouba/alger
Emploi : etudiante
Date d'inscription : 21/09/2008
 |  Sujet: La transcription du matériel génétique Sujet: La transcription du matériel génétique 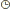 Mar 2 Nov - 22:02 Mar 2 Nov - 22:02 | |
| Chez les eucaryotes, le transcrit primaire d'ARNm est complété par une queue et une tête, c'est la coiffe de guanosine méthylé.
- La transcription est un processus biologique ubiquitaire qui consiste, au niveau de la cellule, en la copie des régions dites codantes de l'ADN en molécules d'ARN. En effet, si la molécule d'ADN est le support universel de l'information génétique, ce sont les molécules d'ARN qui sont reconnues par la machinerie de traduction en séquences protéiques.
- L'enzyme qui catalyse cette réaction de transcription est nommée ARN polymérase. Il en existe plusieurs types intervenant dans la transcription de plusieurs types d'ARN (messager, ribosomique, de transfert, etc. ) L'ARN polymérase reconnaît et se fixe sur une région spécifique de l'ADN, localisée en amont d'une région codante d'un gène : le site promoteur.
- Chez les eucaryotes, le transcrit primaire d'ARNm est complété par une queue (polyadénine) et une tête (riche en guanine), c'est la coiffe de guanosine méthylé.
La molécule d'ARN directement synthétisée à partir du modèle ADN reste dans le noyau et est traitée par un complexe enzymatique. Ce mécanisme se nomme l'épissage : certaines séquences nommées introns sont excisées, les exons restant se relient ensuite entre eux. Il peut y avoir un mécanisme d'épissage alternatif, augmentant ainsi le nombre de possibilités d'ARN messager mature. L'ARN produit est plus court, passe dans le cytoplasme et devient un ARNm ou ARN messager mature.
L'ARNm est alors traduit en protéine à partir des acides aminés en présence des ribosomes et des ARN de transfert (ARNt). Ce mécanisme se nomme la traduction.
Chez les procaryotes
La transcription a lieu dans le cytoplasme bactérien puisque c'est là que se trouve l'ADN (chromosome ou plasmide) Elle a lieu en 3 étapes différentes : l'initiation, l'élongation et la terminaison. L'initiation s'effectue au niveau du promoteur, immédiatement en amont de la séquence codante. L'ARN polymérase bactérienne est constituée de 4 sous unités : α, β, β'et σ. La sous unité σ reconnaît une séquence consensus dans le promoteur du gène (classiquement, la "Pribnow-Box" [TATAAT], se fixe sur l'ADN et recrute les autres parties du complexe enzymatique. Une fois l'ARN polymérase mise en place, la sous unité σ se détache du complexe et β'assure la liaison à l'ADN : c'est le complexe fermé. L'ARN polymérase déroule ensuite la double hélice sur 17 paires de bases environ : c'est le complexe ouvert. La phase d'élongation peut commencer en remplaçant le départ de la sous unité σ par une Nus A. Le complexe enzymatique synthétise alors le brin d'ARN complémentaire du brin codant. La terminaison de la transcrition se fait au niveau du terminateur, localisé après la séquence codante. Des séquences d'ADN riches en paires G-C avec 3 liaisons hydrogènes, par conséquent plus fortes, ralentissent la progression de l'ARN pol, et d'autre part la formation d'une boucle en épingle à cheveux entre 2 régions complémentaires de l'ARN bloquent l'enzyme. Enfin une protéine de terminaison (Rho) libére la molécule d'ARN. L'ARNm ainsi produit est directement utilisable par la machinerie cellulaire pour être traduit. Comme la transcription et la traduction se déroulent dans le même compartiment, il est même envisageable que des ribosomes commencent à traduire un ARNm avant que sa transcription soit terminée.
Chez les eucaryotes
De nombreuses différences existent comparé à la transcription chez les procaryotes. La transcription se déroule dans le noyau cellulaire. La chromatine doit au préalable avoir été décompactée (euchromatine) pour permettre à la machinerie protéique d'accéder à l'ADN. Contrairement aux procaryotes, l'ARN produit par la transcription n'est pas directement utilisable par les ribosomes et devra subir plusieurs étapes de maturation post-transcriptionnelle.
Enfin il existe 3 types d'ARN polymérase différentes au lieu de l'unique pour les procaryotes. Mais seules, ces ARN polymérases ne peuvent rien faire et elles doivent être accompagnées de facteurs de transcription généraux (protéines). Ces facteurs s'appellent TFI, TFII, TFIII... ce qui forme un complexe protéique constitué de 8 à 14 sous unités.
Comme auparavant on retrouve les 3 phases différentes dans le processus de transcription à commencer par l'initiation ; L'équivalent chez les eucaryotes de la «boîte de Pribnow» est la «boîte TATA» localisé à peu près 30 paires de bases avant l'origine de transcription, celle ci joue un rôle prépondérant dans la mesure où elle va fixer l'ARN polymérase II. Deux autres «boîtes» font aussi partie des séquences consensus parmi elles : la boîte CAAT qui est un site modulateur de la transcription et la boîte GC. Enfin on trouve une région dite «Enhancer» qui peut stimuler la transcription à plusieurs centaines de paires de bases du lieu de la transcription.
Par la suite les différents facteurs de transcription viennent s'assembler sur ce «noyau». La TFII A vient stabiliser le complexe TFII D-ADN, la TFII B et TFII E se lient à l'ADN, la TFII F qui est une hélicase ATP-dépendante et enfin l'ARN polymérase II. Cependant ce complexe ne peut initier la transcription qu'à une faible fréquence. Des facteurs de transition supplémentaires doivent intervenir. Parmi eux le CTF (NF1) se lie à la boîte CAAT, le Sp 1 se lie aux boîtes GC : ce sont des trans-activateurs. D'autre part, les séquences «enhancers» «cis activateurs» seront elles-mêmes activées par des protéines activatrices et par une boucle sur elle-même de l'ADN ces mêmes protéines vont entrer en contact avec la boîte TATA.
L'initiation par l'ARN polymérase débute par la protéine TFII D qui est elle-même constituée de la protéine de liaison TBP qui va se fixer sur la boîte TATA ce qui va former le cœur du complexe d'initiation.
L'élongation peut alors commencer ; l'élément central de l'élongation est la phosphorylation du domaine CTD (Carboxy Terminal Domain), domaine spécifique de la sous unité de 220 kDa de l'ARN polymérase II. Celle-ci est riche en sérine et en thréonine qui sont deux acides aminés pouvant être phoshorylés sur leur groupement hydroxyle. La phosphorylation par TFII H (du domaine CTD) qui est une protéine Kinase en présence d'ATP va déplacer l'ARN polymérase jusqu'au lieu d'origine de la transcription. L'addition séquentielle des ribonucléotides peut alors démarrer.
La terminaison est assurée par des signaux spécifiques dont le signal de polyadénylation AATAAA. la polymérase continue sa transcription légèrement après ce motif puis est libérée sous l'effet de divers facteurs. La transcription elle-même est terminée mais l'ARN obtenu n'est pas fonctiuonnel pour tout autant et doit subir 3 étapes de maturation. L'ARN est clivé au niveau du signal de polyadénylation et une polymérase spécifique (la polyA Polymérase ou PAP) ajoute de nombreux résidus Adénine (50 chez les levures, 200 chez les eucaryotes supérieurs) : c'est la queue polyA, principale à la stabilité de l'ARN. Il est à noter que cette partie de l'ARN n'est pas codée dans l'ADN sous forme de polyT.
À l'autre extrémité 5', l'addition d'une coiffe méthylguanosine est indispensable pour la reconnaissance par les ribosomes lors de l'étape de traduction. Il faut malgré tout noter que les SnRNA qui sont aussi synthétisés par l'ARN pol II ont une coiffe mais ne passent pas dans les ribosomes pour être traduits !
L'ARN obtenu n'est pas toujours prêt à être traduit et doit subir une dernière étape de maturation post-transcriptionnelle. En effet, l'ADN chez les eucaryotes possède des séquences codantes (Exons) et des séquences non codantes (Introns). Seuls les exons participent par conséquent à la synthèse des protéines. L'ARN des eucaryotes est en premier lieu produit sous forme de pré-ARNm qui contient toute la séquence du gène (introns exons). Il subit ensuite une opération d'épissage : un complexe nucléoprotéique (le spliceosome) reconnaît les introns et les élimine (l'épissage permet en outre d'obtenir différentes protéines à partir d'un même gène en sélectionnant quels exons seront conservés : c'est l'épissage alternatif).
Les ARN 45S futurs 28 ; 18 ; 5.8S
Le gène 45S se trouve sur des séquences spécifiques de 5 chromosomes (13, 14, 15, 21, 22), nommées organisateur nucléolaire. Sur chacun de ces chromosomes il existe à peu près 40 copies du gène. C'est la transcription intense de cet ADN en ARN qui crée le nucléole. Sur l'ADN 45S, la reconnaissance de la TATA ne peut se faire directement. Il faut passer par la reconnaissance préalable de deux zones consensus : le promoteur central aux environs de 1 et UCE (upstream control element) plus en amont. UBFI reconnaît UCE ce qui entraîne la courbure de l'ADN. Ainsi la TATA peut être reconnue par la TBP et les 3 TAFI de SLI. L'ARN polymeraseI est alors recrutée et peut démarrer directement la synthèse. Elle n'a pas de CTD et son simple recrutement permettrait de la faire démarrer. Cette synthèse s'arrête brusquement au niveau de séquences de terminaison.
Les gènes de classe III
Il s'agit des ARNt, de l'ARN 5S et des SCARN dispersés dans le génome. Les mécanismes changent dans les 3 cas. Leur particularité principale est de posséder des promoteurs internes c'est-à-dire en aval de 1.
- les ARNt
Possèdent deux séquences consensus : les boîtes A et B. Elles sont reconnues par TF IIIC. TF IIIB avec sa TBP et ses 2 TAF III peut alors se fixer. L'ARN polymérase III est recrutée. TF IIIC s'en va et la synthèse peut alors commencer. Possèdent une boîte C reconnue par TF IIIA. TF IIIC et B peuvent alors se fixer. L'ARN polymérase est recrutée et Les TF IIIA et C s'en vont. La synthèse peut démarrer. Divers facteurs intermédiaires permettent la fixation des TAF III et de TBP de TF IIIB. La polymérase est recrutée et la synthèse démarre. La Transcription InverseLa transcription inverse (ou rétrotranscription) est la réaction inverse de la transcription. C'est la synthèse d'un brin d'ADN à partir d'une matrice ARN grâce à une ADN polymérase ARN dépendante ou encore ou rétrotranscriptase (le terme anglais reverse transcriptase est quelquefois aussi utilisé). | |
|
aissou khaoukha
Nombre de messages : 2
Age : 32
Localisation : bejaia darguina
Emploi : étudiante
Date d'inscription : 15/11/2013
| | Sujet: Re: La transcription du matériel génétique Ven 22 Nov - 12:19 | |
| | |
|

